Izadi Jakintza»Izadi jakintza
Zelula prokariotoen ugalketa
LABURPENA: Zelula prokariotoen ugalketa, oro har, asexuala izaten da, zatiketa bitar
bidezkoa; hau da, guraso zelulatik bi zelula sortzen dira, informazio genetiko bera dutenak.
Dena dela, zenbaitetan, zelula prokariotoek geneak trukatzen dituzte elkarrekin. Trukatze hori
hiru prozesuren bidez egiten dute: konjugazioa, eraldatzea eta transdukzioa. Transdukzioan
birus batek egiten du bitartekari lana.
Sarrera
Hau da zelularen ugalketaren urrezko araua: belaunaldi berriko zelula guztiek jaso behar dute zelula sortzailearen DNA kopia bat. Zelula berriak ez dira behar bezala hazten, baldin eta ez badituzte DNA-ren argibide guztiak jasotzen.
Bakterioak sexugabe ugal daitezke zeharreko fisio bikoitzaren bidez, edo, gutxiagotan, gai genetikoaren trukearen bidez berkonbinazio genetikoa. Trukea hiru mekanismoren bidez gertatzen da: eraldatzea, konjugazioa eta transdukzioa.
Bakterioek ahalmen handia dute sexugabe ugaltzeko, gauza baitira 20 minutuz behin zatitzeko; beraz, ugalketa prozesuan inongo interferentziarik ez bada, esaterako elikagaien amaitzea edo hondakin gaien bidezko inhibizioa, sei bat ordutan 250.000 bakterio sor ditzake bakterio batek.
Bakterio askok esporak eratzeko gaitasuna dute; hori, ordea, ez da ugalketa mota bat, baizik eta egoera kaltegarriei aurre egiteko modua. Bakterioko espora bat sortzen da; beraz, zelula kopurua ez da aldatzen. Dena dela, espora horiek ez dira nahastu behar onddoen edo zenbait landareren esporekin, azken horiek ugalketa prozesuaren emaitza baitira.
Zeharreko fisio bitarra
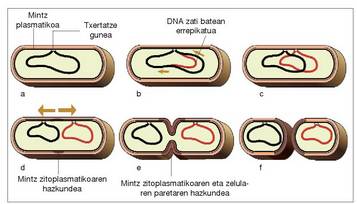
Zelularen ugalketa metodo sinpleena da: zelula bakar batetik bi zelula ume sortzen dira. Zelula ume bakoitza jatorrizkoaren berdin-berdina da; beraz, jatorrizko osagarri guztiak bikoiztu eta bi zelula umeen artean banatu behar dira. Prozesu horrek hiru aldi ditu: -DNA-ren bikoiztea -DNA-ren banatzea -Zelula umeak bereizteko pareta bat osatzea Mailaz maila gertatzen den gertaera segida horri zelularen zatitze zikloa esaten zaio.Mesosomek zuzentzen dute prozesu horretan
gertatzen den DNA-ren bikoiztea eta
bakterio berriak banatuko dituen mintz
plasmatikoaren osatzea.
Zelula prokariotoetan, mintzik gabeko
molekula da DNA, eta horixe da, hain
zuzen, bikoiztu behar dena. DNA molekula,
bikoiztu aurretik, plasma mintzari finkatua
dago mesosoma batean (1A irudia). Ondoren,
DNA bi norabidetan bikoizten hasten
da (1B irudia). Behin bikoizketa amaitu
denean, DNA molekula berria plasma mintzari
finkatuta gelditzen da, jatorrizko DNA
finkatuta zegoen gunetik hurbil (1C irudia).
Plasma mintza hazi egiten da, eta, horren
ondorioz, poliki-poliki banandu egiten dira
bi DNA molekulak (1D irudia). Bi molekulen
arteko gunean, zelularen pareta berria
eta mintz berria sortzen dira; azkenik, bi
zelula umeen banaketa gertatzen da (1E,F
irudia).
Gerta daiteke osatu berria den zelularen
pareta erabat ez zatitzea, eta, horren ondorioz,
bakterio koloniak osatzea (estreptokokoak,
diplokokoak, etab.).
380
Berkonbinazio genetikoa zelula prokariotoetan
Monera erreinuko izakiek ez dute gametoen fusio bidezko sexu ugalketa konplexurik izaten; hala ere, zenbaitetan, hartu-eman genetikoa izaten da (berkonbinazio genetikoa).
Trukatze hori animalia mota desberdinen artean ere gerta daiteke (prozedura parasexualak).
Prozesu horien ondorioz, bi genoma bereizietan dauden osagai genetikoak banako bakar batean batzen dira. Beraz, genotipo mota berriak sor daitezke. Prozedura hori egokia da giro berrietara egokitzeko; monera erreinuko izakien artean, ordea, gutxitan gertatzen da.Prokariotoen berkonbinazio genetikoan,
zelula emaile bateko DNA-ren zati bat, exogenote
deitua, txertatzen da zelula hartzaile
batean, eta zati hori zelula hartzailearen
genoman edo endogenotean sartzen da. Bi
zati horiek elkartzen direnean zigoto partziala
sortzen da. Zelula prokariotoetan, eukariotoetan
ez bezala, ez dira benetako zelula
diploideak sortzen, emailearen gai genetikoaren
parte bat besterik ez baitzaio ematen
hartzaileari, eta parte hori diploide bihurtzen
da zelula hartzailearen gai genetikoaren
zati bati buruz; zati hori zelula emailetik
jasotako zatiaren antzekoa da.
Hiru metodo daude DNA zatia zelula
emailetik hartzailera eramateko: eraldatzea,
transdukzioa eta konjugazioa.
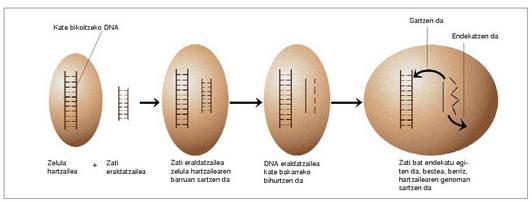
Eraldatzea
Pneumokokoan (Streptococcus pneumoniae) aurkitua. Prozesu hau bakterio jenero batzuetan gertatzen da bakarrik (Bacillus, Neisseria, Pseudomonas, Hemophilus). Bakterio zelula-hartzaile batean beste bakteriozelula puskatu batetik datorren kate bikoitzeko
DNA askea sartzen da; DNA puska
hori zelula hartzailearen barruan sartzen da,
dagokion DNA segmentuan. Sartu ondoren,
kate bakarreko bihurtzen da kate
bikoitzeko puska. Ondoren zatietako bat
endekatu egiten da, eta bestea, berriz, hartzailearen
genoman sartzen da.
Zelula guztiak ez dira eraldagarriak izaten,
batzuek bakarrik dute ahalmen hori. Zelula
eraldagarri horiei zelula konpetente esaten
zaie, hau da, gauza dira DNA eraldagarria
zurgatu eta ezaugarri berriak barneratzeko.Batetik bestera igarotzen den DNA zatia oso
txikia izaten da.
Konjugazioa
Prozesu hau Escherichia coli-n aurkitu zen.
Berkonbinazio genetikoa gerta dadin, ezinbestekoa da bi zelulen artean kontaktua edo konjugazioa izatea. Bi zelulatako bat DNA emaile gisa edo ar gisa aritzen da, eta hartzaile edo eme gisa bestea. Eraldaketan eta transdukzioan ez bezala, konjugazioan genoma zati handiak transmititu daitezke.
Geneetan zehazten da emaile edo hartzaile gisa aritzeko ahalmena. Bakterio bat ar izatea birusaren antzeko agente kutsatzaile bati dagokio, eta faktore sexual, episoma edo F (emankortasun) esaten zaio (3.. rudia). Faktore horrek emeak kutsatu eta ar bihur ditzake. F duten zelulei eta transmititzen dutenei F + (ar) esaten zaie, eta F ez dutenei F- (eme). Bada beste bakterio emaile mota bat, Hfr (berkonbinatzeko maiztasun garaia) esaten zaiona, izan ere, bere genomaren zati handi bat maiztasun handiaz igortzeko ahalmena baitu. F +-ak, ordea, genoma zati txikia besterik ez du igortzen. Bai F + bai F faktorea libre daude zitoplasman; Hfr bakterio motatan, ordea,bakterioko DNA-ren barruan dago F faktorea.
F faktorea F +-tik Fra igor daiteke, eta
orduan F + bihurtzen da; aldi berean, F + bat
Hfr bihur daiteke, baldin eta F faktorea bakterioaren
genoman sartzen bada.
Konjugazioa lau alditan gertatzen da:
- zelula emailea eta hartzailea parekatzea.
Prozesu honetan garrantzi handikoak dira ile
sexualak, izan ere, gai genetikoaren trukatzea
errazten baitute. F + zelulek eta Hfr-ek ileak
izaten dituzte, F- zelulek, berriz, ez dute ilerik.- DNA zati kate-bakar bat zelula emailetik
hartzailera aldatzea. DNA zati hori, edo
emailean sintetizatzen da, Farekin lotura
gertatzen den unean bertan, edo emaileak
berea du lehendik, lotura gertatu aurretik.
Katea Fan sartu, eta berehala barneratzen
da zelula horren genoman.
- emailearen DNA-ren eta hartzailearen
DNA-ren arteko berrantolaketa.
- fenotipo berriaren sorrera.
Konjugazioa zenbait bakterio jeneroren
laboratorioko haztegietan frogatu ahal izanda, hala nola, Escherichia, Shigella, Salmonella,
Pseudomonas, Serratia eta Vibrio generoetan.

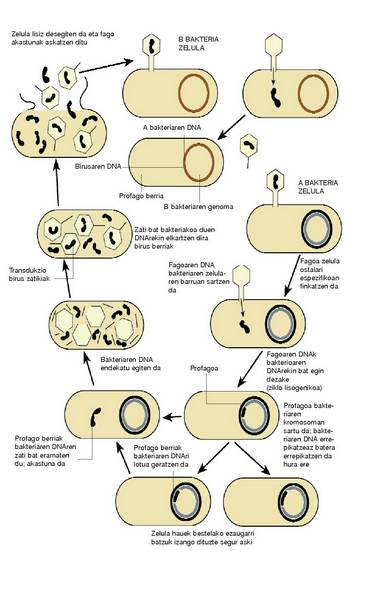
Transdukzioa
Prozesu honetan igortzaile bat birus bakteriofagoa edo fagoa behar izaten da; azken bakterio kutsatutik hartutako DNA zatiak garraiatzen ditu igortzaile horrek.
Birus transduktore esaten zaio. Beraz, berrantolaketa genetikoa gertatuko bada, bakteriobakterio harremana ez da nahitaezkoa.
Ziklo lisogenikoan (4. irudia), birus baten DNA zati bat (profagoa) bakterioaren genoman sartzen da, eta bakterioaren zelulen zatiketa gertatu ahala gai genetiko hori errepikatu egiten da. Profagoa ziklo litikoan sartzeak zenbait ondorio dakartza: profagoa bakterioaren DNA-tik bereiztean, bakterioaren gai genetikoaren zati bat berarekin eramatea; bakterioa zelulan eta horren lisian ugaltzea; zelula berriak kutsatzea. Horrek dakar profagoa aurretiko bakterio baten gai genetikoa daramana, beste bakterio baten genoman sartzea, eta horren ondorioz genoma hori diploide partziala izango da lehen bakterio kutsatutik zekarren gai genetikoari buruz.
Birus zatiki transduktorea batetik akastuna da, izan ere, DNA zati bat galdu baitu (bakterioaren DNA-ri itsatsita gelditu dena), eta, bestetik, ez du gaitasunik bakterio ostalarian ziklo litiko bat eragiteko.
Bi transdukzio mota daude: orokorra eta mugatua, edo espezializatua.
Transdukzio orokorrean bakterioko gene guztiek fagora eransteko aukera dute, izan ere, bakterioaren DNA katearen edozein puntutan sar baitaiteke fagoaren DNA; birusa ziklo litikoan sartzen denean eta bakterioaren DNA-tik bereizten denean, birus helduaren zatikiaren parte bihurtzen da DNA hori, birusaren DNA izan ordez, edo izateaz gainera. Hala ere, era horretan sortutako zatiki transduktoreek duten fagoaren DNA kopurua ez da atzemateko adinakoa; aitzitik, fago baten kapsidearen barruan bakterioaren DNA zati bat dutela dirudi.
Transdukzio mugatuan, profagoaren txertatze gunearen alboko geneak bakarrik sartzen dira fagoan, honek bakterioaren DNA uzten duenean. Txertatze gune hori ia beti bera denez gero, transduzitutako geneak gene jakin batzuk dira (mugatuak). Zatiki transdukzio eragilean dagoen bakterioaren DNA fagoaren DNA-ren zati handi bati lotuta dago, eta birusaren zenbait generen ordezko da. Birus zatiki berri horien DNAk nahasian ditu birusaren eta bakterioaren DNA, eta errepika badaitezke ere, akastunak dira, izan ere, birusak bere geneetako batzuk galdu baititu. Birus zatiki berri horiek gaitasuna dute bakterio ostalari batera jaso dituzten bakterio geneak transduzitzeko; hala ere, akastunak direnez gero, ezin ekin diote prozesu litikoari.