Izadi Jakintza»Izadi jakintza
Bilakaeraren teoria sintetikoa

1940. urte ingurukoa da bilakaeraren teoria sintetikoa, Darwinen ideiak eta genetikaren
aurkikuntzak bateratzen dituena. Teoria honen abiapuntua Darwinen hastapen
bat da; hastapen honek dioenez, populazio biologikoetan ageri diren bariazio genetikoei
eragiten diela-eta, hautaketa naturala da bilakaeraren motorra. Ingurunearen ezaugarrien
arabera, era askotakoa izan daiteke hautaketa horren eragina.
Bariazioaren eta hautaketaren ondorioz sortzen dira mota berriak (espeziazioa),
aurreko motetatik abiatuta. Prozesu hori gertatzeko, beharrezko da ugalketa hesiak
agertzea, mota baten baitan organismo talde bakartuak sor daitezen. Mekanismo
bakartzaile horiek geografia espeziazioaren ereduan oso mantso, bi alditan, agertzen
dira, eta bat-batean berriz espeziazio lasterraren ereduan.Mende honen erdia arte ia, Darwinen
hautaketa naturalaren teoria zen zientzialarien
artean nagusia. Teoria horrek ordea
eragozpen handiak zituen bariazioak eta
bariazio horiek nola heredatzen diren esplikatzeko.
Mendelen legeen berraurkikuntzaren
ondoren (1900) eta De Vriesek mutazioak
aurkituta, auzitan jarri zen hautaketa
naturala aski ote zenik bilakaeraren prozesu
nagusi gisa. Genetikaren aurkikuntza berriak
ez ziruditen bat zetozenik Darwinen ideiekin:
genetika bariazio kualitatiboez mintzo zen
(bariazio bat-bateko eta zehatzez), eta pixkana-pixkana
pilatzen diren bariazio kuantitatibo
txikiez berriz Darwin zaleak.
Bi joeren arteko eztabaidak hamarralditan
luzatu ziren, garbi-garbi frogatu zen arte
ezaugarri kuantitatiboek ere (pisuak, neurriak,
etab.) Mendelen legeak betetzen zituztela,
baina, gene bakar batek eraginak izan
gabe, gene bakar baten baino gehiagoren
mendeko izanik (herentzia faktore-anitza
edo kuantitatiboa). Era horretan hasi ziren
genetika eta Darwinen teoria elkarrengana
hurbiltzen; azkenean, 1937an, Dobzhanskyk,
Genetika eta moten jatorria argitaratu
zuenean, bilakaeraren teoria sintetikoaren
oinarriak ipini zituen. Hurrengo
hamarraldian behin betiko garatu zen teoria
hori, hainbat zientzilariren lan bikainari
esker: Mayr, Simpson, Huxley, etab.
Gaur egun teoria sintetikoak du onespen
zabalena, baina teoria berriak ere agertu
dira, hala oreka etendunaren teoria, eta mutazio
neutroena.





Teoria sintetikoa
Neodarwinismo ere esaten zaio. Darwinen teoria nagusiak jasotzen ditu, genetikak ekarritako ezagutza berrien argitara berriro aztertu eta osaturik. Bilakaeraren teoria dagaur egun nagusia zientzialarien artean.
Teoria honek bi zutabe ditu: populazioen
bariazio genetikoa, eta ugaritasun horretan
eragiten duen hautaketa naturala.
Bariazioa da bilakaeraren oinarria. Bi
bariazio mota dira: giroak sortutako bariazioak
(Lamarckek zioen organoen erabiltze
eta ez erabiltze hura), eta jatorri genetikoa
duten bariazioak. Azken horiek dira, genetikoak
alegia, bilakaerari ardura zaizkionak,
horiek baitira ondorengoei transmitituko
zaizkienak.
Mutazio genetikoak dira bariazio heredagarriaren
lehen iturria. Horien gainean beste
mekanismo batzuek eragiten dute, bariazio
hori asko areagotzen dutenak. Mekanismohorien artean batzuk aipa daitezke, hala nola
migrazioak, noraez genetikoa, etab., eta,
batez ere, sexu bidezko ugaltzea, zeinak,
meiosiaren eta ernalketaren bitartez, diren
forma guztiak konbinatzen baititu, eta
mugagabe zabaltzen ugaritasuna. Indar
horiek normalean zoriaren arabera jokatzen
dute, ez baitago indar jakinik mutazioak
zentzu baterantz edo besterantz bideratuko
dituztenik.
Hautaketa naturala
Kontzeptua
Bai Darwinen teoriak, bai bilakaeraren
egungo teoria sintetikoak, hautaketa naturala
jotzen dute izaki biziek denboran zehar izan
duten aldaketa edo bilakaeraren arrazoi nagusietakotzat.
Populazio batean diren genotipoen
ugaltze tasa desberdin gisa definitzen da
hautaketa naturala. Desberdintasunaren arrakasta
da. Gehien ugalduko direnak ingurugiro
horretara hobekien egokitutakoak izango
dira (?egokienak?, esan ohi zuen Darwinek),
eta proportzio handiagoetan transmitituko
dizkiote beren aleloak hurrengo belaunaldiari.
Horren ondorioz, belaunaldi gazteak gene
osakera desberdina izango du, eta, hala, bilakaera
gertatuko da.
Ikusi dugu lehen aldaketaren abiapuntua
mutazioak direla, hau da, alelo berriak
agertzea. Mutante berri bat agertzen denean
hautaketak haren gainean eragiten du, alelo
berri horren arabera. Alelo gainartzailea
bada, laster agertuko da genotipoan, eta
gauzatu ahal izango da hautaketaren eragina,
alde nahiz kontra. Adibide baterako,alelo gainartzaile batek eragindako eritasun
hilgarri batean kasuan, desagertu egingo
dira alelo hori duten banako guztiak ondorengo
belaunaldietan. Baina, atzerakoia baldin
bada, alelo horren homozigotoak besterik
ez dira desagertuko.
Aurreko adibidetik ondorioztatzen den
beste puntu bat da zer garrantzi izan dezakeen
alelo berri horrek organismoaren bizitza
osoan. Milaka gene horien arteko bat besterik
ez bada, hau da, ez badu eraginik funtzio
garrantzitsuen gainean, ez dira asko igerriko
agian alelo horren ondorioak fenotipoaren
osotasunean, eta hautaketan ere ez da asko
antzemango. Badirudi horrela jokatzen
duela gehienbat hautaketak, pixkana-pixkana
eragiten diola fenotipo osoari, eta mailakako
bilakaera bat eragiten duela.
Alelo baten eraginkortasuna bere organismoa
bizi den ingurugiroaren araberakoa izaten
da. Ingurugiroaren baldintzetara hobekien
egokitzeko gai direnak izaten dira alelo
eraginkorrenak. Hala, kolore zuria oso
baliotsua izan daiteke Artikoan bizi diren
ugaztunentzat (hartz zuriaren kasua). Ingurugiroaz
mintzatzean, osagai desberdinak
hartu behar dira kontuan, biotikoak nahiz
abiotikoak. Batzuetan janaria izango da faktore
garrantzitsua, tenperatura besteetan,
edo argia, harrapariak, etab.
Hautaketa naturalaren indarra hautaketa
presio esapidearen bitartez adieraz daiteke.
Alelo bat hautaketa presiope handian egoteak
esan nahi du hautaketa hori ez duela oso bere
aldekoa, hau da, alelo hori duten banakoak oso
gutxi ugaltzen direla, edo, gauza bera dena, oso
eraginkortasun biologiko apala dutela.
Kontzeptua
Bai Darwinen teoriak, bai bilakaeraren egungo teoria sintetikoak, hautaketa naturala jotzen dute izaki biziek denboran zehar izan duten aldaketa edo bilakaeraren arrazoi nagusietakotzat.
Populazio batean diren genotipoen ugaltze tasa desberdin gisa definitzen da hautaketa naturala. Desberdintasunaren arrakasta da. Gehien ugalduko direnak ingurugiro horretara hobekien egokitutakoak izango dira (?egokienak?, esan ohi zuen Darwinek), eta proportzio handiagoetan transmitituko dizkiote beren aleloak hurrengo belaunaldiari.
Horren ondorioz, belaunaldi gazteak gene osakera desberdina izango du, eta, hala, bilakaera gertatuko da.
Ikusi dugu lehen aldaketaren abiapuntua mutazioak direla, hau da, alelo berriak agertzea. Mutante berri bat agertzen denean hautaketak haren gainean eragiten du, alelo berri horren arabera. Alelo gainartzailea bada, laster agertuko da genotipoan, eta gauzatu ahal izango da hautaketaren eragina, alde nahiz kontra. Adibide baterako,alelo gainartzaile batek eragindako eritasun
hilgarri batean kasuan, desagertu egingo
dira alelo hori duten banako guztiak ondorengo
belaunaldietan. Baina, atzerakoia baldin
bada, alelo horren homozigotoak besterik
ez dira desagertuko.
Aurreko adibidetik ondorioztatzen den
beste puntu bat da zer garrantzi izan dezakeen
alelo berri horrek organismoaren bizitza
osoan. Milaka gene horien arteko bat besterik
ez bada, hau da, ez badu eraginik funtzio
garrantzitsuen gainean, ez dira asko igerriko
agian alelo horren ondorioak fenotipoaren
osotasunean, eta hautaketan ere ez da asko
antzemango. Badirudi horrela jokatzen
duela gehienbat hautaketak, pixkana-pixkana
eragiten diola fenotipo osoari, eta mailakako
bilakaera bat eragiten duela.
Alelo baten eraginkortasuna bere organismoa
bizi den ingurugiroaren araberakoa izaten
da. Ingurugiroaren baldintzetara hobekien
egokitzeko gai direnak izaten dira alelo
eraginkorrenak. Hala, kolore zuria oso
baliotsua izan daiteke Artikoan bizi diren
ugaztunentzat (hartz zuriaren kasua). Ingurugiroaz
mintzatzean, osagai desberdinak
hartu behar dira kontuan, biotikoak nahiz
abiotikoak. Batzuetan janaria izango da faktore
garrantzitsua, tenperatura besteetan,
edo argia, harrapariak, etab.
Hautaketa naturalaren indarra hautaketa
presio esapidearen bitartez adieraz daiteke.
Alelo bat hautaketa presiope handian egoteak
esan nahi du hautaketa hori ez duela oso bere
aldekoa, hau da, alelo hori duten banakoak oso
gutxi ugaltzen direla, edo, gauza bera dena, oso
eraginkortasun biologiko apala dutela.
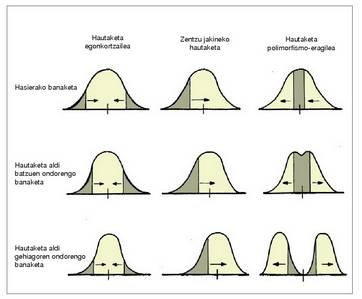
Hautaketa motak
Era askotakoak izan daitezke hautaketak populazio baten gainean izaten dituen ondorioak. Hori dela-eta, hainbat motatan sailkatu da hautaketa. Ikus ditzagun horietako batzuk.
Hautaketa egonkortzailea
Bilakaera aldaketarik ez dakarrena da.
Banako gehienak berentzako egokiena den ingurugiroan bizi diren populazioetan aurkitzen dugu. Egoera horietan, hautaketaren presioak populazioaren muturreko banakoen gainean baizik ez du eraginik; tarteko
balioak dituzten banakoek berriz arrakasta
handiagoa izaten dute.
Kasu hau hobeto ulertuko da ikerketatik
ateratako adibide bat ikusiz. 1930. urte
inguruan zenbait gizatalderen jaio ondorengo
hilkortasuna aztertu zenean, ikusi zen
jaiotzean tarteko pisuak zituzten haurrek
bizi iraupen luzeagoa zutela gutxiegi edo
gehiegi pisatzen zutenek baino. Gaur egungo
sendagintzaren arretarik gabe, gora egingo
luke tarteko pisuko fenotipoen maiztasunak.
Zentzu jakineko hautaketa
Ingurugiroko baldintzak aldatzen direnean gertatzen da. Halakoetan erraz gerta daiteke ondoen egokitutako fenotipoa aurreko hoberen berbera ez izatea, eta banaketaren grafikoan hoberen horretatik urrun egotea. Horren ondorioz, gora egingo du hoberen berriaren iraupen itxaropenak, eta banaketa aldatu egingo da, fenotipo hori banaketaren erdian geratzen den arte. Orduan gertatuko da aurreko hautaketa egonkortzailea. Hautaketa motahonetan ere, jakina, izan da bilakaerarik.
Zenbaitetan ez da beharrezkoa izaten ingurugiroa
aldatzea hautaketa mota hau gerta
dadin; aski izan daiteke egokitze hobetzeren
bat ekarriko duen bariazioren bat zoriz
agertzea.
Dudarik gabe, Biston betularia pipien
kasua da (?industria melanismo? gisa ezagutzen
dena) zentzu jakineko hautaketaren
kasurik aipatuena. Joan den mendearen
bukaera aldea arte Biston betularia motako
pipi guztiak gris kolorekoak ziren, arrasto
beltzekin. Garai horretan, hego beltzeko
pipiak hasi ziren agertzen Manchesterren.
Forma berri hori zabalduz joan zen pixkanapixkana,
industria esparruetan nagusia izatera
heldu zen arte, landareak kutsadurak
belztuak zeuden lekuetan alegia.
Hautaketaren eragilea, kasu horretan,
txoriak ziren, pipiak jaten baitituzte. Pipi
beltzak askoz hobeto ezkuta daitezke zuhaitz
kutsatuen enbor beltzetan, industria zabaldu
aurretik ez bezala. Fenotipo onenak ilunaren
aldera jo zuen, hilkortasuna apalagoa
baitzuen, eta, hala, 1895ean, beltzak ziren
Manchesterko pipi guztien %98. Aldi berean,
guztiz kontrakoa zen banaketa industriarik
gabeko nekazaritza eremuetan, forma
beltzik ez baitzegoen ia.
Egoera horrek bere horretan iraun zuen
1960. urte ingurua arte, orduan hasi baitziren
aplikatzen kutsaduraren aurkako
lehen legeak. Neurri horiek berriro aldatuzuten ingurugiroa, eta zuhaitzen enborrek
antzinako koloreak hartu zituzten berriro.
Egoera horretan, fenotipo argia zen komenigarriena,
eta berriz ere gertatu zen hautaketa,
pipi argiak areagotu eta ilunak
desagertarazteko joerakoa. Gaur egun,
pipi beltz gutxi dago berriro industria eremuetan.
Ezagutzen dira zentzu jakineko hautaketaren
beste adibide asko ere, hala mixomatosiaren
birusa gainditu duten untxiak, pozoinari
buru egin dioten arratoiak, edo
antibiotiko askori buruz inmune diren
mikrobio familiak. Hautaketa artifiziala,
bestalde, asko erabiltzen dute animalia edo
landare hazleek.
Hautaketa polimorfismo-eragilea
Ingurugiroak, aldi berean, bi fenotipo edo gehiagoren alde egiten duenean gertatzen da hauteketa mota hau; uniformetasun gutxi duten lekuetan gertatzen da, ingurugiroaren egoera toki batetik bestera aldatzen denean.
Bi forma abantailatsuenak, edo gehiago, gertatuko dira hautatuak, eta populazioko forma nagusi bihurtuko dira; era horretan, talde desberdinak sortuko dira populazio horretan, hau da, polimorfismoa gertatuko da.
Behin baino gehiagotan aipatu izan da mea hondakinak dauden eremuetan hazten den belar mota baten adibidea. Metal batzuek, hala kobreak edo berunak, asko kutsatu dituzten lurrak dauden lekuetan, mota horren aldaera bat nagusitu da. Horren aldamenean,kutsatu gabeko lur esparruetan,
beste bat hazten da. Normal-normal gurutzatzen
badira ere, lur mota bakoitzean hartara
egokitutako aldaera besterik ez da hazten.
Adibide horretatik ondoriozta daitekeenez,
lehen populazio batek, inguruaren parte
baten eraldatzearen ondorioz, bi azpimotaren
bereiztea ekarri du.
Asko eztabaidatzen den giza polimorfismoaren
kasu bat larruazalaren kolorearena
da. Ezaugarri hori, genetikan ikusi den bezala,
genetikaren eta ingurugiroko faktoreen
mendekoa da. Munduko parte desberdinetako
jatorrizko gizataldeen larruazal koloreen
banaketa aztertzen denean, badirudi
ezaugarri horri ere eragin diotela hautaketa
naturalaren faktoreek; irrada ultrabioletaren
kontrako babesa eta D bitaminaren beharra
dira hautaketa horren faktore nagusiak.
Mota berrien sorrera
Atal honetan, galdera honi heltzea komeni da lehen-lehenik: zer da mota? Zientziari ez zaio erraza gertatu erantzuna aurkitzea.
Historian zehar, bi oztopo handi izan dira motaren kontzeptuaren azterketan: zientzialari askoren ustea zen motarik ez zela benetan, aitzitik, kategoria arbitrario hutsa zela, komeni zelako ematen zitzaiona izaki talde bati, morfologia definizioa egin ahal izateko; hala, kanpoko itxuraren arabera sailkatzen ziren motaren ezaugarriak.
1940-1950. urteetan egin zen motaren definizio modernoa: ?Mota bat antzeko ezaugarriak dituen banako talde batek osatzen du; banako horiek elkarrekin gurutzatuz ugal daitezke, eta ondorengo emankorrak sortzen dituzte. Izaki multzo horrek gene ondare komuna du, beste mota batena ez bezalakoa.
Aitzitik, bi izaki talde mota desberdinekoak dira, elkarren ondoan bizi direla, ezin direnean gurutzatu, edo, gurutzatzen badira, ezin badute ondorengo emankorrik eman?.
Ikusten denez, kontzeptu hori motaren bi alderdi biologikoetan oinarritzen da: ugaltze bakartasunean eta gene ondarean, eta, hori dela-eta, motaren kontzeptu biologikoa esan ohi zaio. Azpimarratu beharra dago definizio honek sexu ugalketa duten motentzat balio duela ?halakoxeak dira gehienak?, ez ordea sexu gabekoentzat, bakterioentzat adibidez.
Nolanahi ere, mota batean, normalean, populazio bat baino gehiago izaten da, esparru desberdinetan kokatua, zeinek bere gene ondarea duela. Aparte daudela, bakoitzak bere bilakera berezia izan dezake, eta, denborarekin, elkarren arteko aldeak areagotu daitezke.
Espeziazioa edo aurrekoetatik abiatzen diren mota berrien sorrera da bilakaeraren gertaera nagusia. Gaurko teoria sintetikoak prozesu hori hautaketa naturalaren oso antzera agertzen du, orain genetikari dagozkion xehetasunak erantsi zaizkion arren,
Darwinen teoria osatzen dutenak.
Mota kontzeptutik beretik datorrenez,
banako talde bat bere motatik bereizteko eta
beste mota bat, berria, bihurtzeko, hesiak edo
mekanismo biologikoak agertu behar dute
batez ere, hesi horiek aldenduko baitituzte
jatorrizko motatik. Mekanismo bakartzaile
hauen agertzea oinarri-oinarrizko baldintza
da espeziazioa gerta dadin, eta, hedaduraz,
baita bilakaera izan dadin ere. Hainbat eratako
mekanismoak deskribatu dira; horietako
batzuk ondorengo laukian jasotzen dira.
Populazio bereko izaki bizien arteko hesi
hauek nola eratu diren agertzerakoan ez
datoz bat ikerlariak. Teoria desberdinak proposatu
diren arren, hurrengo alditan labur
daiteke prozesua.Lehen aldian, gelditu egiten da populazio bereko bi talderen arteko gene trukea, bai banaketa fisikoz (geografia hesiak), bai talderen bati eragin dion aldaketa genetiko baten ondorioz. Lehenengoa da izadian gehien ikusi den kausa.
Gene trukerik ez badago, gero eta desberdinagoak egingo dira bi taldeak. Horren esplikazioa da bariazioa dakarten faktoreek, batez ere hautaketa naturalak, baina orobat mutazioak, noraezak, sexu berkonbinazioak, etab., modu desberdinean eragin ohi diotela talde bakoitzari. Denbora puska baten ondoren, ugalketa bakartasun postzigotikoa hasiko da agertzen. Banaketak irauten baldin badu, bi mota desberdin izatera iritsi daitezke.
Gerta daiteke halaber bi moten arteko geografia hesia desagertzea ere bakartzea hasi berria denean, eta, hala, berriz ere ugaltzeko gai izango dira. Hibridoak guztiz bideragarriak baldin badira (hibrido ez direnak bezala), ugalketa bakartze hasi berri hori ezereztu egingo da, eta hasierako populazioa bere horretan geratuko da. Baina hibridoak ez badaude besteak bezain egokituak, bigarren aldian sartuko dira. Aldi horretan, areagotu egingo da ugalketa bakartasuna (prezigotikoa agertuko da), eta, azkenean, mota desberdin bihurtuko dira.
Lehen aldia, oro har, luzea izaten da, aurrerago ikusiko diren zenbait salbuespen kenduta. Bestalde, askotan ez da bigarrenik gertatzen, adibidez, taldeak uharte desberdinetara emigratu duten kasuetan.
Espeziazio alopatrikoa edo geografikoa
Hauxe da teoria sintetikoaren aldezle gehienek onartzen duten eredua. Prozesu motela eta mailakakoa izaten da. Eremu zabalean bizi den mota bat baldin bada, erraz bereiziko dira geografia hesiek (mendi, ibai, itsaso eta abarrek) banaturiko populazioetan. Populazio horiek giro desberdinen eraginpean biziko dira, eta horrekegokitze mota desberdinak sorraraziko ditu
organismoetan, eremuen arabera. Hesiek
populazioen arteko gene trukea eragotziko
dute, eta aldeak gero eta handiagoak izango
dira. Milaka edo milioika urte igaro ondoren,
populazio horietako batzuetan ugalketa
bakartzeko mekanismoak agertuko dira,
mota desberdinen sorrera ekar dezaketenak.
Espeziazio eredu hori, bere bi aldiekin,
Ameriketako Drosophila mota batzuk aztertzean
frogatu da. Lehen aldia D. willistoni
motan gertatzen da; Ande mendiek banaturiko
bi populazio ditu: D. w. willistoni, mendien
ekialdean, eta D. w. quechua, mendebalean.
Bi taldeak laboratorioan gurutzatu
direnean, hibridoak partez antzuak dira;
bakartasun postzigotikoko mekanismoak
agertu dira beraz.
Ameriketako leku horretan bertan bada
beste mota bat, D. paulistorum, espeziazioaren
bigarren aldia bizi duten hainbat populazio
talde dituena. Talde horietako batzuk
eremu berean bizi dira, hau da, sinpatrikoak
dira, baina ez dira ugaltzen, sexu jokabide
desberdinak baitituzte (bakartasun prezigotikoa).
Mota sortu berritzat jo daitezke horiek.
Espeziazio lasterra edo kuantikoa
Sinpatrikoa ere deitu izan zaio, lurralde berean bizi diren populazioetan artean gertatzen baita zenbaitetan, hau da, banaketa geografikorik gabe.
Bakartasun mekanismo batzuk gutxi-asko bat-batean agertzen dira, eta mota bereko taldeen arteko gurutzaketak eragozten dituzte. Mekanismo horiek banako bakar batean ager daitezke, eta banako hori gai izan da mota berriko populazio bat sortzeko.
Kasu horietan ez da izaten espeziazioaren lehen aldirik, edo, izatekotan, oso laburra da. Hori da espeziazio mota bakunena, De Vriesek eta Mendelen lehen jarraizaileek defenditzen zutena.
Era honetako kasu asko ezagutzen dira, gene edo kromosoma mutazioak pairatu dituzten landareenak batez ere; mutazio horiek hibridoen bizi iraupena murriztu dute, eta, pixkana, populazioaren gainerako kideetatik bakartuko dira. Dudarik gabe, era honetako prozesurik argiena poliploideena da.
Poliploidia angipospermo mota askoren eragiletzat hartzen den mutazioa da, angipospermo landuen eragiletzat batez ere.
Landare horietako gehienak tetraploideak izaten dira. Animalietan, berriz, oso gutxitan gertatzen da, sexualitate berezia dutenetan bakarrik hain zuzen ere: partenogenesia, hermafroditismoa, etab.
Poliploidian, bi edo hiru belaunalditan sor daiteke mota berri bat; alopoliploidia izaten da gehienetan. Gogoratu behar da AAeta BB izendatuko ditugun bi kromosoma
sail dituzten bi mota diploide gurutzatuta,
ondorengo hibrido diploide bat emango
dutela (AB), antzua. Batzuetan hibrido
horrek bere kromosomak autobikoizten
ditu, eta AABB tetraploidea bihurtzen da,
eta haren ondorengoek autoernalketa bidez
landare emankorrak dituen populazio bat
sor dezakete. Tetraploide hori, jakina, ez da
gai izaten bere arbasoen tankerako motekin
gurutzatzeko, eta, hortaz, mota berritzat
hartzen da. Espeziazio mota honi mota artekoa
ere esaten zaio. Autopoliploideen kasua
ikusi da jada mutazioetan.
Bilakaeraren beste teoria batzuk



Oreka etendunak edo aldizkakoak
Neodarwinismoan, mota berrien agertzea oso aldi luzeetan zehar gertaturiko aldaketa txikien pilaketaren emaitza da. Baina, 1972an, beste zientzialari batzuek, Eldredgek eta Gouldek adibidez, bestelako teoria bat proposatu zuten, oreka etendunen edo aldizkakoen teoria deritzana; teoria horren arabera bi gertaera guztiz desberdin hurrenez hurren gertatzen direnean bilakatzen dira biologia taldeak: badira alde batetik aldi egonkor luzeak, motak aldatzen ez direnak; bestalde, tarteka eten gogorrak izaten dira, gutxi irauten dutenak, baina mota berriak emango dituzten aldaketa bortitzak gertaurteetanzehar, eta desagertu egiten diren.
Hurrena, beste mota batzuk agertzen dira,
guztiz desberdinak, bitarteko urratsik gabe,
eta horietan ere gauza bera gertatzen da:
denboraldi batez aldagabe mantendu, eta
desagertu egiten dira.
Hainbat kasu aipatu dira ?saltokako? bilakaera
mota honen alde, hala adibidez hominidoen
burezurraren edukia bat-batean handitzea,
mota batetik bestera igarotzean (mota
beraren baitangehitzerik antzeman ez dela),
edo oso mutazio gutxiren eraginez sortu ahal
izan diren mota berrien kasua (giza mota eta
txinpantzea bereizten dituzten kromosoma
mutazioen kasua). Duela gutxi, iraganeko
moten iraungialdi handien aurkikuntza
ikusgarriek ere (Burgessko kanbrikoko fauna
Kanadan, Palezoikoaren amaierakoa, etab.)
teoria honen alde egiten dute.
Gaur egun, zientzialari askoren aburuz,
bateragarriak dira teoria hori eta sintetikoa;
izan ere, zenbait motaren agerpenean erritmo
biziagoa izan daitekeela baizik ez baitu proposatzen,
eta hori aurreko teoriak ere onartzen
zuen, Darwinek berak ere aipatu baitzuen.
Mutazio neutroak
1968an Ohta eta Kimura zientzialariek neutralismoaren teoria proposatu zuten; teoria horren arabera, mutazio gehienak neutroak izaten dira, hau da, ez dute ondoriorik organismoan, ez baitiote ez abantailarik ez desabantailarik ekartzen egokitzeari. Populazio bateko alelo neutroen maiztasuna aldatuz joango da belaunaldietan zehar, modu guztiz aleatorioz, hautaketa naturalak hartan parte hartu gabe. Denboraren igarotzearekin, gene neutro batzuk ugaltzen edo desagertzen hasten badira, asko aldatuko da populazio horren osakera genetikoa, mota berri bat sortzen den arte. Beraz, teoria honetan zoriak du eginkizun garrantzitsuena; bilakaerarik gertatzen da, nahiz eta hautaketa naturalak jokatu ez.
Teoria berri honen oinarria froga biokiomikoetan dago batez ere. Gaur egungo DNAren eta proteinen sintesiaren ezagutzak frogatu du molekulen aldakortasuna uste zena baino garrantzitsuagoa zela, eta bariazio neutroak ere asko izaten direla. Mutazio genetikoen ondorioak aztertzen direnean, ikusten da nola gene baten nukleotido bati bakarrik eragiten dioten aldaketa batzuek lor dezaketen gene horri dagokion proteinan aminoazido baten lekua beste batek hartzea. Aldaketa txiki horrek proteinaren jardunean izan dezakeen eragina garrantzitsua izan daiteke, baina gerta daiteke halaber aminoazido bakar baten aldaketa horrek proteinaren jardunean batere ondoriorik ez sortzea. Horrez gainera, nukleotido baten aldaketak ez du aminoazido baten ordezkatzea zertan eragin behar, kodoi berriakaurrekoaren esanahi bera balu gertatuko litzatekeen
bezala. Pentsa dezagun, adibidez,
badela GCA kodoia GCC bihurtzen duen
mutazio bat: zer ondorio izango lukehorrek? Hala bada, badira, izan, mutazio
neutroak, eta mutazioak beren horretan
irautea edo desagertzea zoriaren araberakoa
da soil-soilik.
